EVOLUCIÓ
| lloc: | Cursos IOC - Batxillerat |
| Curs: | Biologia II (Bloc 2) ~ gener 2020 |
| Llibre: | EVOLUCIÓ |
| Imprès per: | Usuari convidat |
| Data: | dimecres, 26 de juny 2024, 12:15 |
Descripció
.
Taula de continguts
- 1. Concepte d'espècie i fet evolutiu
- 2. Evolució: evidències
- 3. Teories de l'evolució
- 4. Genètica de poblacions i evolució
- 5. Especiació
- 6. Història de la Terra
- 7. Resum sobre lamarquisme, darwinisme i neodarwinisme
- 8. Exemple de selecció natural i tolerància a la lactosa
- 9. Un altre exemple de selecció natural : coevolució
- 10. Errades freqüents
1. Concepte d'espècie i fet evolutiu
El concepte d’espècie des d’un punt de vista morfològic fa referència a un grup d’organismes que comparteixen unes característiques morfològiques determinades. És una concepció utilitzada en la vida quotidiana i que ja feien servir els primers naturalistes. Però el dimorfisme sexual (és a dir, les diferències entre mascles i femelles) i el polimorfisme, ja presents en algunes espècies, creen dificultats en aquesta definició.

Femella i mascle de faisà (Phasianus colchicus)
Però, des d’un punt de vista biològic, una espècie és un grup d’organismes que es poden encreuar entre ells i donar descendència fèrtil. És la definició més acceptada per la comunitat científica, però cal destacar:
• Només es pot aplicar a organismes amb reproducció creuada (com els animals i els vegetals).
• No es pot aplicar als organismes que només es reprodueixen asexualment (com els bacteris).
• Tampoc no es pot aplicar als organismes fòssils, dels quals no sabem amb quins altres organismes es podien encreuar i amb quins no.
• En la pràctica, no es pot aplicar intensivament a totes les poblacions.
El concepte biològic, quan es pot aplicar, és el més utilitzat. En els altres casos es fa servir el concepte morfològic.
El fet de l’evolució
En biologia, l’evolució és el procés segons el qual la diversitat d’éssers vius s’ha original per canvis transmesos a la descendència. L’evolució implica canvis en les poblacions i en les espècies al llarg de les generacions, però no en els individus; és a dir, el desenvolupament, el creixement i els canvis durant la vida d’un organisme no es consideren evolució.
Cal tenir en compte que l’evolució és un fet, com demostren un bon nombre d’evidències. El que la comunitat científica discuteix actualment no és si existeix o no l’evolució, sinó quina teoria pot explicar millor com es produeix. De fet, els qui avui dia encara neguen l’evolució ho fan de manera pseudocientífica o clarament dogmàtica.
2.1. semblances anatòmiques
Les semblances anatòmiques
Si es comparen animals diferents, s’hi troben les evidències següents:
• Òrgans homòlegs. òrgans amb forma externa i, fins i tot, funció diferents , però amb la mateixa estructura interna , perquè tenen el mateix origen embriològic i evolutiu . En són exemples les extremitats anteriors dels rèptils, les aus i els mamífers
https://www.deviantart.com/amadeublasco/art/Forelimb-bones-homology-454625204
Les homologies en l’extremitat anterior dels vertebrats tetràpodes indiquen un origen evolutiu comú.
• Òrgans anàlegs. Òrgans amb una estructura i un origen embrionari diferents, però que exerceixen la mateixa funció. No tenen el
mateix origen evolutiu. En són exemples les ales dels insectes i les aus.
• Òrgans vestigials. Òrgans que no són útils per a l’animal, però que en els seus avantpassats es trobaven molt més desenvolupats i feien una funció útil. En són exemples l’apèndix vermiforme i el còccix humans, i petits ossos de la cintura pelviana de les serps i les balenes.
2.2. semblances embrionàries
Les semblances embrionàries
Quan comparem embrions de diferents espècies de vertebrats, observem els aspectes següents:
• Els primers estadis embriològics de tots els vertebrats són molt semblants. Després es diferencien seguint aproximadament la jerarquia taxonòmica.
• Tots els vertebrats, incloent-hi els terrestres, passen per un estadi embriològic en el qual tenen fenedures branquials.
• Els embrions humans, com els dels altres vertebrats, passen per una etapa durant la qual tenen cua.
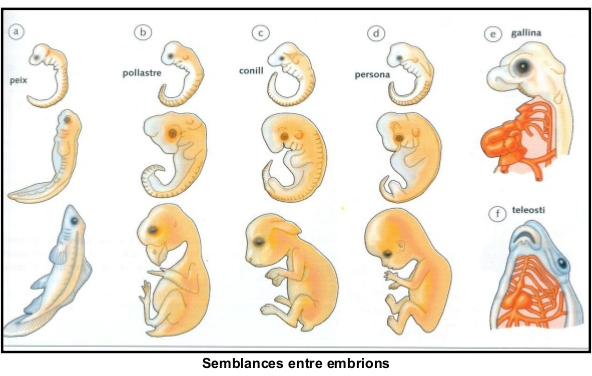
Llibre de Conceptes bàsics. Teide.
2.3. els fòssils
Els fòssils són restes d’organismes d’altres temps, o de la seva activitat, que per causes naturals s’han conservat en les roques. La paleontologia és la ciència que estudia els fòssils.
Molts fòssils corresponen a ossos, ous, closques o altres parts dures petrificades, però també són fòssils els organismes conservats en gel, resines o asfalts. Igualment són fòssils les petjades, les marques o altres restes de l’activitat dels éssers vius conservades a les roques.
A partir dels fòssils, els paleontòlegs poden extreure molta informació de l’organisme que els va deixar i deduir aspectes sobre el seu entorn i, en alguns casos, fins i tot poden arribar a la reconstrucció completa del cos de l’organisme.
L’antiguitat dels fòssils es pot determinar:
• directament, utilitzant mètodes de datació radioactiva;
• indirectament, tenint en compte la disposició relativa dels estrats que els contenen.
El registre fòssil, el conjunt de fòssils que es coneix, és incomplet. Això és conseqüència, entre d’altres, dels motius següents:
• El fet que un organisme deixi fòssils és poc freqüent.
• La fossilització és un procés llarg i difícil, que es dóna sobretot en sediments dipositats en el medi aquàtic.
• La fossilització de parts toves d’organismes encara és més difícil, perquè aquestes parts es deterioren en poc temps.
Aquesta dificultat és fonamental en el cas dels organismes formats només per parts toves.
• La major part dels fòssils que hi ha actualment estan enterrats a l’interior de les roques, en l’escorça terrestre.
Malgrat aquestes dificultats, els paleontòlegs han observat que:
• Cada era, període o època en la història de la Terra presenta un conjunt de fòssils característics, els fòssils guia.
• Els éssers vius han anat canviant al llarg del temps. Cada espècie apareix en un moment determinat, es conserva durant
períodes més o menys llargs de temps i s’acaba extingint.
• Els grups d’éssers vius apareixen en un moment determinat, a partir d’un grup anterior. Per exemple, els primers rèptils van sorgir fa uns 325 milions d’anys (a partir d’un grup d’amfibis), i els primers mamífers, fa uns 210 milions d’anys (a partir d’un grup de rèptils).
• Com més separades estan dues èpoques en el temps, més diferents són els seus fòssils; per exemple, els fòssils del cretaci (fa 100 milions d’anys) són molt més semblants als del juràssic (fa 150 milions d’anys) que als del carbonífer (fa 300 milions d’anys).
• En molts casos s’han trobat successions d’espècies semblants, esglaonades en el temps, que en conjunt mostren una evolució ramificada. Un dels casos més divulgats és el dels avantpassats dels cavalls.
També s’han trobat formes de transició entre grups d’organismes diferents. Un dels més famosos és el fòssil d’Archaeopteryx, que presenta característiques intermèdies entre els rèptils i les aus.

2.4. biogeografia
La biogeografia és la ciència que estudia la distribució geogràfica de les espècies vives.
Cada continent i cada illa mostren una flora i una fauna particulars, amb espècies diferents de les d’altres llocs, encara que amb característiques adaptatives similars; per exemple, a l’Amèrica del Sud i l’Àfrica, amb climes similars, hi
viuen espècies diferents, com el jaguar i el lleopard.
Això és conseqüència del fet que les espècies existeixen únicament en els ambients amb condicions favorables que han estat colonitzats pels seus avantpassats; per exemple, els avantpassats dels lleopards van arribar a l’Àfrica, però no a l’Amèrica del Sud.
En molts arxipèlags, com ara les Galápagos o les illes Hawaii, hi ha molt pocs grups d’organismes vius, cadascun dels quals té una gran diversitat d’espècies endèmiques (que només es troben allà). Això s’explica perquè els pocs organismes que van arribar a aquests arxipèlags els van colonitzar i van evolucionar de manera diferent a cada illa, i així es va originar la gran diversitat d’espècies actuals.

2.5. selecció artificial
Els animals domèstics i les plantes cultivades provenen d’animals i plantes salvatges, criats pels éssers humans des de fa uns pocs milers d’anys. Moltes vegades, aquests animals i plantes tenen aspectes força diferents dels seus parents salvatges, com passa en els gossos i els llops.
En algunes espècies domèstiques hi ha varietats d’aspecte tan diferent com un gos pastor alemany i un gos terrier escocès .
En aquests animals i plantes domèstics, es trien com a reproductors els individus que es consideren millors, els que tenen les característiques més valorades. Per exemple, els bens que donen més llana, les vaques que fan més llet, el blat més productiu o més resistent als factors climàtics, etc.
Amb aquest procés de selecció artificial hem fet evolucionar, en uns pocs milers d’anys, els animals domèstics i les plantes conreades, utilitzant un mecanisme equivalent a la selecció natural.

2.6. proves bioquímiques i moleculars
Universalitat dels mecanismes vitals bàsics
Tots els éssers vius coneguts de la Terra utilitzen els mateixos mecanismes moleculars bàsics, tot i que aquests mecanismes podrien funcionar bé igualment de manera diferent. Això ens indica un origen evolutiu comú.
• Utilitzem DNA com a material genètic.
• Fem servir RNA com a intermediari en l’expressió de la informació genètica.
• Usem les proteïnes com a enzims i per a realitzar nombroses funcions biològiques.
• Construïm les proteïnes amb els mateixos 20 aminoàcids.
• Utilitzem gairebé el mateix codi genètic per llegir la informació genètica.
• Fem servir ATP per transferir energia.
• Utilitzem glucosa com a font d’energia i per construir altres substàncies.
Comparació de cèl·lules i orgànuls
Si es comparen els organismes de la Terra, es pot observar que tots els animals tenim cèl·lules molt similars, i també són semblants les cèl·lules dels vegetals. Tant les cèl·lules dels animals com les dels vegetals són diferents de les cèl·lules dels bacteris.
En aquesta comparació, també es fa evident que les espècies evolutivament properes tenen cromosomes molt similars . En canvi, les espècies evolutivament distants tenen cromosomes molt diferents.
Comparació de seqüències gèniques
Alguns gens i algunes proteïnes són presents en un gran nombre d’éssers vius. Habitualment, cada espècie en presenta la seva pròpia seqüència, semblant a les seqüències d’espècies evolutivament properes i molt més diferent d’altres espècies; per exemple, el gen que codifica la cadena beta de l’hemoglobina en humans té 444 parells de nucleòtids.

llibre de Conceptes bàsics. Editorial Teide
3. Teories de l'evolució
Apunts sobre les primeres teories sobre la història de la vida, neodarwinisme i especiació: teories de l'evolució
Ens interessa sobre tot conèixer les característiques del lamarckisme, del darwinisme, del neodarwinisme ( al punt 7 hi ha un resum) i l'especiació (punt 5)
Vegeu aquest vídeo de 6 min sobre Darwin i l'evolució https://www.youtube.com/watch?v=2qsE2j0jJ-I
4. Genètica de poblacions i evolució
La genètica de poblacions estudia la composició genètica de les poblacions i com varia aquesta composició en funció del temps.
- Caracterització genètica de les poblacions mitjançant les freqüències gèniques i genotípiques
Freqüències al·lèliques
El nombre d’organismes d’una població que posseeixen un determinat al·lel determina la freqüència d’aquest al·lel en la població. Per exemple, si en l’espècie humana la freqüència de l’al·lel dominant per a la producció de pigment a la pell, els ulls i el cabell és del 99%, l’al·lel recessiu responsable de la manca de pigmentació (albinisme) presenta una freqüència de l’1%.
En genètica de poblacions les freqüències al·lèliques s’acostumen a expressar amb decimals:
|
Freqüència al·lel dominant (A) |
0.99 |
|
Freqüència al·lel recessiu (a) |
0.01 |
|
Suma de les freqüències dels dos al·lels |
0.99 + 0.01 = 1 |
S’acostumen a utilitzar els símbols estadístics “p” i “q” per a simbolitzar les freqüències amb les que dos al·lels, l’un dominant (“A” de freq. “p”) i l’altre recessiu (“a” de freq. “q”) apareixen en el pool o reservori de gens d’una població. Per tant:
p + q = 1
-
Càlcul de les freqüències al·lèliques, gèniques i genotípiques
La relació matemàtica entre les freqüències al·lèliques i genotípiques en les poblacions va ser desenvolupada independentment per Hardy i Weinberg i se la coneix com a equilibri de Hardy-Weinberg. Es basa en el principi de que la freqüència dels al·lels recessius i dominants en una població és constant al llarg de les generacions si no intervenen elements modificadors.
Si “p” es la freqüència de l’al·lel “A” i “q” la de l’al·lel “a” les freqüències de les tres combinacions possibles d’aquests dos al·lels seran:
-
A (p)
a (q)
A (p)
AA (p2)
Aa (pq)
a (q)
Aa (pq)
aa (q2)
-
Genotip
Freqüència
genotípica
AA
p2
Aa
2 pq
aa
q2
i es compleix que :
p2 + 2pq + q2 = 1
-
Exemples de càlculs de freqüències al·lèliques i genotípiques
-
Amb dos al·lels autosòmics codominants
Quan dos al·lels són codominants, cada fenotip correspon a un genotip diferent.
Si en una població de N individus una característica està determinada per dos al·lels A i B codominants:
-
D individus seran homozigòtics AA
-
R individus seran homozigòtics BB
-
H individus seran heterozigòtics AB
I a més:
D + R + H = N
Donat que els organismes són diploides el nº total d’al·lels és 2N.
Freqüència de l’al·lel A = p = (2D + H) / 2N = (D + ½ H) / N
Freqüència de l’al·lel B = q = (2R + H) / 2N = (R + ½ H) / N
-
Amb dos al·lels autosòmics, un dominant i un recessiu
Amb dos al·lels, un dominant “A” i un recessiu “a”, el fenotip dominant pot correspondre a dos genotips diferents (AA i Aa); l’únic fenotip del qual podem saber el genotip sense cap dubte és el recessiu “aa”.
En una població de N individus:
-
D seran AA
-
H seran Aa
-
R seran aa
La freqüència del genotip recessiu “aa” serà R/N i aquesta freqüència és igual a q2.
Es pot calcular així la freqüència de l’al·lel recessiu “a” (q) i la de l’al·lel dominant “A” (p = 1 – q).
4.1. Factors que fan variar les freqüències gèniques a les poblacions (agents evolutius)
Hardy i Weinberg van demostrar que la recombinació genètica que té lloc en cada generació no canvia per si mateixa les freqüències gèniques de les poblacions sempre que es donin les 5 condicions següents:
|
|
És difícil que aquestes 5 condicions es donin en una població natural i per tant l’equilibri és impossible i en les poblacions reals les freqüències dels al·lels sempre estan canviant.
Els processos que permeten que les freqüències de les variacions genètiques canviïn de generació en generació són les mutacions, el flux gènic, la deriva genètica i la selecció natural. Tots aquests processos, excepte l’últim, són preadaptatius: tots ells produeixen canvis en les freqüències gèniques independentment de si aquests canvis milloren o redueixen l’adaptació dels organismes al seu ambient. La selecció natural és l’únic que promou l’adaptació i que limita els efectes desorganitzadors dels altres processos.
Mutacions
Una mutació és un canvi en l’estructura o la quantitat de l’ADN d’un organisme. Això produeix un canvi en el genotip que pot ser hereditari. Les mutacions que es donen en les gàmetes són hereditàries, en tant que les que es donen en les cèl·lules somàtiques tan sols es transmeten a les cèl·lules que deriven per mitosi de la cèl·lula mutada.
Es distingeixen dos grans tipus de mutacions: les gèniques són canvis puntuals en l’estructura de l’ADN, en tant que les cromosòmiques afecten a la quantitat o a la distribució de l’ADN.
Les mutacions són l’origen de noves característiques genètiques a les poblacions mitjançant canvis espontanis del material genètic.
Flux gènic o migracions
Són moviments d’organismes o de grups d’organismes d’un hàbitat o localitat a un altre. El flux genètic pot introduir al·lels nous en una població o bé modificar les freqüències dels al·lels ja existents. El seu efecte global és una tendència a reduir les diferències entre poblacions.
Deriva genètica
Es tracta de canvis aleatoris no direccionals de les freqüències gèniques, determinats per la grandària finita de les poblacions o pel seu aïllament. En poblacions petites, fenòmens aleatoris com per exemple la mort prematura per accident de l’únic individu portador d’un determinat al·lel pot eliminar aquest al·lel de la població (un al·lel que tingui una freqüència de l’1% estarà present en un sol individu d’una població de 50, però si la població és de 100.000 individus hi haurà 2000 al·lels repartits entre molts individus; és improbable pensar que per atzar puguin desaparèixer els 2000 al·lels, però si l’únic individu portador mor prematurament o sense deixar descendència, aquest al·lel desapareixerà de la població de 50). De la mateixa manera es pot justificar que un al·lel incrementi molt la seva freqüència simplement per atzar. Un fenomen associat a la deriva genètica és l’anomenat “efecte del fundador” que es dóna quan una població petita es separa d’una més gran i pot no ser del tot representativa de la població d’origen en termes de freqüències al·lèliques (exemples: els Amish de Pennsilvània o la distròfia muscular a Sueca).
Selecció natural
Està determinada per les diferències respecte a la probabilitat de supervivència i de reproducció de cada organisme en el seu ambient. Quan una població creix, alguns factors ambientals poden passar a ser limitants (disponibilitat d’aliments en el cas dels animals, de llum o aigua per a les plantes, ...) Aquest fet comporta que, entre els diferents individus de la població, s’estableixi una competència per a obtenir els recursos necessaris, de tal manera que els organismes millor adaptats sobreviuran i es reproduiran, en tant que els pitjor adaptats probablement moriran sense deixar descendència. Per tant, la selecció natural és el procés que determina quins seran els al·lels que passaran a la següent generació. Segons la teoria sintètica de l’evolució, aquesta és la força principal que actua per canviar les freqüències dels al·lels, és a dir, és el motor principal de l’evolució.
La selecció natural té lloc sempre que es verifiquin les següents condicions:
-
Reproducció: els organismes han de reproduir-se per formar una nova generació.
-
Herència: els descendents han de ser semblants als progenitors.
-
Variabilitat: la població ha de presentar variabilitat genètica.
-
A causa d’aquesta variabilitat, els individus de la població han de tenir una eficàcia biològica diferent (“eficàcia biològica” probabilitat que té un individu per sobreviure i reproduir-se)
Aquestes condicions poden il·lustrar-se amb l’exemple de la Biston betularia : les arnes es reprodueixen, la coloració és hereditària, existeix una variabilitat genètica respecte del model de coloració i les diferents formes tenen diferent eficàcia biològica en funció de l’ambient (les formes fosques o melàniques tenen més probabilitat de sobreviure i reproduir-se en ambients pol·lucionats).

Vegem l’evolució per selecció natural d’una població de mosques de la fruita.
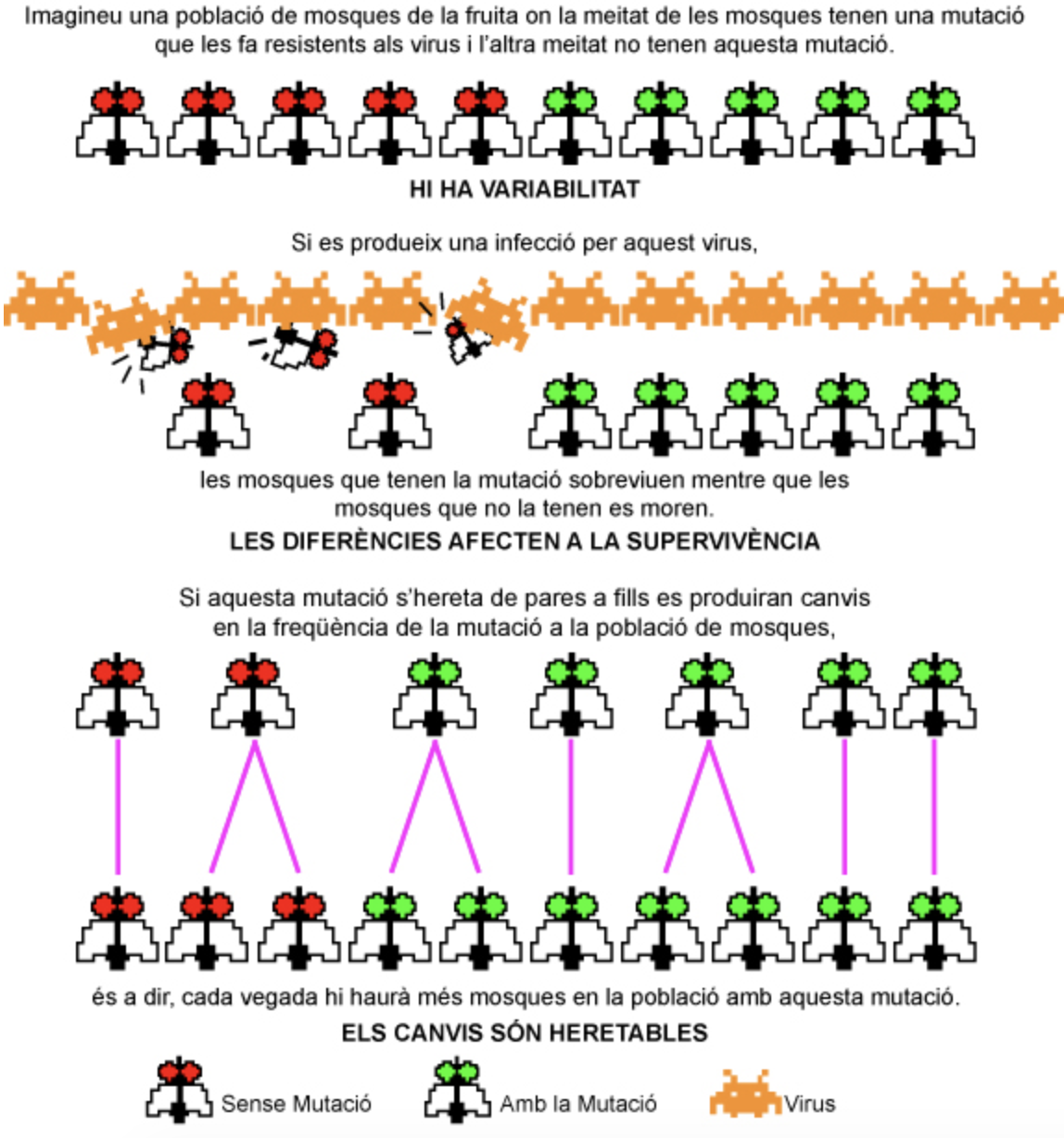
Es donen aquestes tres condicions:
- variabilitat
- diferències en la reproducció i/o en la supervivència
- herència
per això pot actuar la selecció natural que permet als organismes adaptar-se a nous ambients.
4.2. Exemples exercicis càlcul freqüències gèniques
1) En una espècie de vespes, el gen M determina l’amplada normal de l’abdomen i el seu al·lel recessiu m origina un abdomen més estret. Una població d’aquestes vespes està formada per 120 individus homozigòtics dominants, 300 heterozigòtics i 80 homozigòtics recessius. Calculeu les freqüències genotípiques, gèniques i fenotípiques.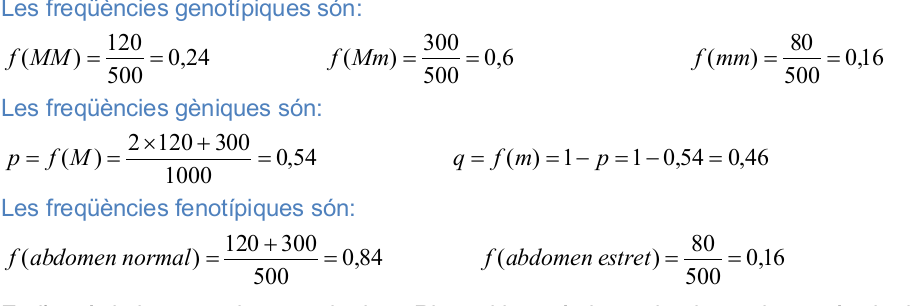
2) En l’espècie humana, la sang de tipus Rh positiu està determinada per la presència de l’al·lel dominant Rh + i el tipus Rh negatiu es determinat per l’al·lel recessiu Rh - . En una població, que suposem que està equilibrada, el 9% dels individus és Rh negatiu. Calculeu les freqüències dels al·lels i dels genotips.
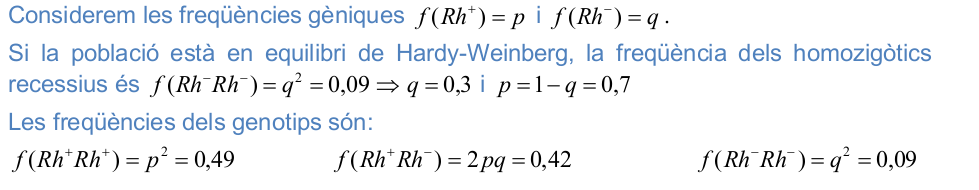
3) En el cargol terrestre Cepaea nemoralis el gen dominant R determina el color marró de la closca i el seu al·lel recessiu r origina closques de color groc. Una població d’aquesta espècie en equilibri de Hardy-Weinberg està formada per 80 individus de color marró i 160 de color groc. Quines són les freqüències gèniques i genotípiques?
Nombre total d’individus: 80 + 160 = 240
Considerem les freqüències gèniques: f(R) = p i f(r) = q
Si la població està en equilibri, f(groc) = q2 = 160/240 ; q = = 0,82
Les freqüències gèniques són q = 0,82 i p = 1 – q = 0,18
Les freqüències genotípiques són:
f(RR) = p2 = 0,182 = 0,032
f(Rr) = 2pq = 0,295
f(rr) = q2 = 0,67
5. Especiació
Vegeu aquest vídeo sobre l'especiació
https://www.youtube.com/watch?v=1hHlvvcAkzE
L'especiació és el procés evolutiu pel qual s'originen noves espècies biològiques. Una espècie no apareix per una mutació que condueix a l'aparició d'un individu d'un tipus diferent. Les espècies s'individualitzen a partir de poblacions que pertanyen a una espècie d'origen (excepte en el cas de certs vegetals). L'especiació resulta de la selecció natural i/o de la deriva genètica, que són els motors de l'evolució. El concepte d'especiació ha estat essencialment desenvolupat per Ernst Mayr.
Existeixen dues formes d'especiació natural basant-se en la
premissa que l'especiació es produeix per la isolació geogràfica de les
poblacions: Especiació al·lopàtrica i simpàtrica. També es pot induir l'especiació artificialment.
Vegeu aquesta animació sobre especiació en llangardaixos:
https://www.youtube.com/watch?time_continue=46&v=EmtIofdeUbc
Especiació natural
Totes les formes d'especiació natural s'han produït en el curs de l'evolució, i encara és un tema de debat la relativa importància de cada mecanisme en la formació de la biodiversistat.
Hi ha diferents opinions sobre el ritme de successos d'especiació que ocorren en el temps geològic. Alguns biòlegs evolutius mantenen que l'especiació s'ha mantingut relativament constant al llarg del temps, mentre que alguns paleontòlegs com Niles Eldredge i Stephen Jay Gould apunten que les espècies es mantenen sense canvis durant llargs períodes de temps, i que l'especiació succeeix només en intervals relativament curts de temps, opinió coneguda com a equilibri puntuat.
Especiació al·lopàtrica
També anomenada especiació geogràfica, és l'especiació gradual que es produeix quan una espècie ocupa una gran àrea geogràfica que no permet que els individus que estiguin molt allunyats puguin creuar-se, a causa de barreres geogràfiques com mars, muntanyes o deserts. La separació espacial de dues poblacions d'una espècie durant un llarg període dóna lloc a l'aparició de novetats evolutives en una o en les dues poblacions degut al fet que el medi ambient és diferent en les diferents zones geogràfiques; es deté el flux genètic entre poblacions. Existeixen dos grans patrons d'especiació al·lopàtrica proposats per Ernst Mayr: l'especiació vicariant i la peripàtrica.
- Especiació vicariant Es produeix quan dues poblacions diferents se separen mitjançant barreres originades per canvis geològics o climàtics impenetrables. Això provoca la fragmentació de l'hàbitat i s'impedeix el flux genètic i es produeix una 'acumulació de diferències tant genètiques com morfològiques. També es pot produir la separació de la població per l'extinció de poblacions intermèdies.
- Especiació peripàtrica: Implica la separació d'una població menor d'una altra més gran, per exemple, quan es colonitza un nou hàbitat. Amb el temps es produeix una divergència genètica i/o morfològica que derivarà en una altra espècie. Aquests individus que migren a un lloc diferent i distant on no existeixen altres membres de la seva espècie i estableixen una nova es coneix com l'efecte fundador.

- Observacions
Existeix la tendència en petites i aïllades poblacions que es produeixin trets característics poc freqüents, s'ha observat en moltes circumstàncies, incloent-hi el nanisme insular i els canvis radicals en cadenes d'illes, com les illes Komodo i les Galàpagos, aquestes últimes han vist néixer la teoria de l'evolució amb les observacions de Charles Darwin. Un dels exemples més famosos d'especiació al·lopàtica és el pinsà de Darwin de les illes Galàpagos.
Especiació simpàtrica
Aquest tipus d'especiació es dóna entre espècies estretament relacionades que conviuen en una mateixa àrea de distribució i que s'especialitzen a ocupar diferents nínxols dins el mateix entorn en què viuen. En aquests casos la selecció natural juga un rol crucial en la divergència de les poblacions. Els mecanismes de l'especiació simpàtrica poden ser:
- Especiació per autopoliploidia És l'especiació espontània en la qual intervé una sola espècie que sofreix una alteració de la meiosi, amb un augment dels cromosomes poliploidia. Solament són viables els múltiples de dos en el nombre de cromosomes (4n, 6n, 8n) i és molt freqüent en els vegetals amb l'aparició d'espècies de gran grandària.
- Especiació per al·lopoliploidia És l'altra especiació espontània en la qual intervenen dues espècies semblants amb el mateix nombre de cromosomes donant lloc a una descendència estèril perquè els cromosomes no són homòlegs. Si durant la gametogènesi de la descendència es produís alguna mutació en la meiosi donant lloc a una poliploidia, s'obtindrien gàmetes amb nombre de cromosomes diploide que només es podran fecundar amb altres híbrids que hagin sofert la mateixa mutació o amb si mateixos.
Aquest tipus d'especiació es pot produir per les causes següents:
- Aïllament ecològic: dintre d'una mateixa zona geogràfica poden existir diferents hàbitats caracteritzats per diferències de temperatura, llum, humitat, etc. que dificultin l'aparellament
- Aïllament estacional: els organismes poden madurar sexualment en diferents estacions o hores del dia.
- Aïllament etològic: es basa en diferències de comportament durant el seguici i l'aparellament com a senyals d'atracció que si fallen provoquen la fugida o l'atac.
- Aïllament mecànic: la còpula és de vegades impossible entre individus de diferents espècies, ja sigui per la grandària incompatible dels seus genitals, o per variacions en l'estructura floral.
- Aïllament sexual: són mecanismes que impedeixen la còpula o la fecundació com les diferències morfològiques dels òrgans reproductors o dels gàmetes
- Aïllament genètic: es produeix en els cromosomes i per tant en la informació genètica. Pot ser de dos tipus:
- Esterilitat dels híbrids: quan dues espècies distintes s'aparellen, la descendència pot ser viable, però estèril.
- Debilitat dels híbrids: quan la descendència de dues espècies distintes no és viable i són eliminats abans d'arribar a la maduresa sexual per selecció natural.


Especiació artificial
Artificialment s'han creat noves espècies a partir de la cria d'animals domestics, encara que les dades inicials i els mètodes d'iniciació de les noves espècies no estan clars. Per exemple, l'ovella domèstica es va crear per hibridació i jo no és possible la reproducció viable amb el mufló, l'espècie de la qual descendeix.[2] Algun tipus de ramat domèstic, per altra banda, es pot considerar com a la mateixa espècie d'algunes varietats salvatges de guineus, iac, etc., ja que encara produeixen descendència fèrtil.[3]
La creació d'una nova espècie en laboratori millor documentada és dels anys 80 del passat segle. William Rice i G.W. Salt van criar mosques de la fruita, Drosophila melanogaster, utilitzant un laberint amb tres opcions d'hàbitat diferents com llum/foscor, i humit/sec. Cada generació es va situar al laberint, i els grups de mosques que sortien de dos de les vuits sortides, s'apartaven per aparellar-les amb altres dels seus respectius grups. Després de 35 generacions, els dos grups i la seva descendència van ser aïllats depenent de les seves preferències d'hàbitat: només s'aparellaven dins les àrees que preferien, i no s'aparellaven amb mosques d'altres àrees
La creació d'una nova espècie en laboratori millor documentada és dels anys 80 del passat segle. William Rice i G.W. Salt van criar mosques de la fruita, Drosophila melanogaster, utilitzant un laberint amb tres opcions d'hàbitat diferents com llum/foscor, i humit/sec. Cada generació es va situar al laberint, i els grups de mosques que sortien de dos de les vuits sortides, s'apartaven per aparellar-les amb altres dels seus respectius grups. Després de 35 generacions, els dos grups i la seva descendència van ser aïllats depenent de les seves preferències d'hàbitat: només s'aparellaven dins les àrees que preferien, i no s'aparellaven amb mosques d'altres àrees. Diane Dodd va poder mostrar especiació al·lopàtrica per reproducció aïllada en mosques de la fruita Drosophila pseudoobscura després de només vuit generacions usant diferent tipus de fruita, midó i maltosa.- Experiment realitzat per Dianne Dodd al 1989 sobre la Drosophila.

6. Història de la Terra
La història de la Terra abasta els aproximadament 4.600 milions d'anys que van des de la formació de la Terra a partir de la nebulosa presolar
fins al present. Aquest article presenta una visió general que resumeix
les teories científiques més acceptades. Per la dificultat de plasmar
períodes de temps molt grans, es farà servir una analogia amb un dia
terrestre, començant fa exactament 4.567 milions d'anys, amb la formació
de la Terra, i acabant ara. Cada segon d'aquest període representa uns 53.000 anys. El Big Bang i l'origen de l'univers,
que es calcula que tingueren lloc fa aproximadament 13.700 milions
d'anys, passarien quasi dos anys abans que comencés a funcionar aquest
rellotge imaginari.
6.1. origen
La Terra es formà com a part del naixement del sistema solar; el que més tard esdevindria el sistema solar existia inicialment com un gran núvol de pols, roques i gasos en rotació. Estava compost d'hidrogen i heli produïts al Big Bang, així com d'elements químics expulsats per les explosions de les supernoves. Aleshores, fa uns 4.600 milions d'anys, es pensa que una estrella propera esdevingué una supernova. L'explosió envià una ona expansiva a través de la nebulosa presolar, causant-ne la implosió. A mesura que el núvol continuava girant, la gravetat i la inèrcia l'aplanaren en un disc protoplanetari, orientat perpendicularment respecte al seu eix de rotació. La majoria de la massa es concentrà al centre i començà a escalfar-se. La implosió d'aquesta matèria donà calor cinètica; com que no es podia escapar, el centre del núvol s'escalfà, permetent la fusió d'hidrogen en heli. El resultat fou un estel T Tauri, el Sol primitiu. Mentrestant, a mesura que la gravetat feia que la matèria es condensés al voltant de les partícules de pols, la resta del disc es començà a separar en anells. Els fragments més petits col·lidien i es tornaven fragments més grans, que esdevenien protoplanetes. Entre ells hi hagué una acumulació de matèria a uns 150 milions de quilòmetres del centre: la Terra. A mesura que el Sol es condensava i s'escalfava, començà a produir-se la fusió nuclear, i el vent solar que en resultà s'endugué la majoria del material del disc que no s'havia condensat en cossos més grans.
La Terra primitiva era molt diferent del món actual. No hi havia oceans i l'atmosfera no tenia oxigen. Rebia l'impacte de planetoides i altres residus de la formació del sistema solar. Aquest bombardeig, juntament amb la calor alliberada pels processos radioactius, la calor residual, i la calor creada per la pressió de la contracció, feia que el planeta es trobés completament fos. Els elements més pesants s'enfonsaren al centre, mentre que els més lleugers pujaren a la superfície, creant les diverses capes de la Terra. L'atmosfera primitiva hauria inclòs el material proper de la nebulosa solar, especialment gasos lleugers com ara l'hidrogen o l'heli, però finalment hauria estat eliminada pel vent solar i la mateixa calor terrestre.
Això canvià quan la Terra tenia un radi d'un 40% de l'actual, i l'atracció gravitatòria permeté la retenció d'una atmosfera amb aigua. Les temperatures s'enfonsaren i l'escorça del planeta s'acumulà sobre una superfície sòlida, amb zones foses per grans impactes. Els impactes a gran escala haurien causat una fusió regional i una diferenciació parcial, portant alguns dels elements més lleugers a la superfície o a l'atmosfera.
La superfície es refredà ràpidament, formant l'escorça terrestre en 150 milions d'anys. Entre fa 4.000 i 3.800 milions d'anys la Terra sofrí un període de bombardeig meteòric. El vapor d'aigua s'escapava de l'escorça mentre que els volcans alliberaven més gasos, formant la segona atmosfera terrestre. Els impactes meteòrics, provinents sobretot del cinturó d'asteroides, aportaren més aigua. El planeta es refredà i es formaren els primers núvols. La pluja originà els oceans fa 3.800 milions d'anys i probablement encara abans; hi ha indicis recents que indiquen que els oceans es podrien haver començat a formar fa 4.200 milions d'anys.
La nova atmosfera contenia probablement amoníac, metà, vapor d'aigua, diòxid de carboni i nitrogen, això com petites quantitats d'altres gasos. L'oxigen quedà atrapat amb hidrogen o en minerals a la superfície. L'activitat volcànica era intensa i, sense una capa d'ozó que l'aturés, la radiació ultraviolada inundava la superfície.
 Les erupcions volcàniques foren comunes a la Terra primitiva
Les erupcions volcàniques foren comunes a la Terra primitiva6.2. Els inicis de la vida
Tot i que se n'han establert els principis generals, no es coneixen detalls sobre l'origen de la vida. Una de les teories proposades és que la vida, o almenys alguns components orgànics, podria haver arribat a la Terra de l'espai, com postula la teoria de la panspèrmia, mentre que d'altres argumenten que és més probable un origen terrestre. De tota manera, els mecanismes causants de l'aparició de la vida serien similars. Si la vida hagués començat a la Terra, la data en què començà és força especulativa; potser fa uns quatre mil milions d'anys. D'alguna manera, en el marc de la química energètica de la Terra primitiva, una molècula (o potser alguna altra cosa) aconseguí l'habilitat de fer còpies de si mateixa - el replicador. No es coneix la naturalesa d'aquesta molècula, ja que la seva funció fou presa fa molt de temps pel replicador actual, l'ADN. Quan feia còpies de si mateix, el replicador no era sempre perfecte; algunes còpies tenien un "error". Si aquest canvi eliminava la capacitat de fer còpies de la molècula, ja no hi hauria més còpies, i la línia "s'extingiria". D'una altra banda, algun canvi rar faria que la molècula es repliqués més ràpidament o millor; aquestes línies esdevindrien més nombroses i "prosperarien". A mesura que les matèries primeres ("el menjar") s'exhaurien, les molècules que podien explotar diferents materials, o potser aturar el progrés d'altres molècules i robar-los els recursos, haurien esdevingut més nombroses.
S'han proposat diferents models per explicar com es podria haver desenvolupat aquest replicador. S'han proposat diferents tipus de replicador, incloent-hi substàncies químiques orgàniques com ara proteïnes modernes, àcids nucleics, fosfolípids, cristalls o fins i tot sistemes quàntics. Actualment no hi ha cap mètode per determinar quin d'aquests models, s'ajusta a l'origen de la vida a la Terra, si és que algun ho fa. Una de les teories més antigues, i que ha estat treballada detalladament, pot servir com a exemple de com podria haver passat això. L'alta energia dels volcans, els llampecs i els raigs ultraviolats podrien haver causat reaccions químiques que haurien produït partícules més complexes a partir de composts simples com ara el metà i l'amoníac. Entre elles hi havia molts dels composts químics orgànics relativament simples que són les peces bàsiques de la vida. A mesura que augmentava la quantitat d'aquesta "sopa orgànica", les diferents molècules començaren a reaccionar entre elles. A vegades es creaven molècules més complexes - potser l'argila serví de base per reunir i concentrar material orgànic.

El replicador
La presència de certes molècules podria accelerar les reaccions químiques. Tot això continuà durant molt de temps, i les reaccions tenien lloc d'una forma més o menys aleatòria, fins que per casualitat aparegué una nova molècula: el replicador. Aquesta molècula tenia l'estranya propietat de causar reaccions químiques que en produïen una còpia, i aquí començà l'evolució. Altres teories proposen un replicador diferent. En qualsevol cas, en un cert punt el paper del replicador fou pres per l'ADN; totes les formes de vida conegudes (exceptuant alguns virus i els prions) utilitzen l'ADN com a replicador, d'una manera gairebé idèntica.
La primera cèl·lula

La vida moderna guarda el seu material replicador ben emmagatzemat dins la membrana cel·lular. És més fàcil entendre l'origen de la membrana cel·lular que el del replicador, ja que les molècules de fosfolípids que componen la membrana cel·lular formen sovint una bicapa de forma espontània quan se les posa dins l'aigua. En certes condicions poden formar-se moltes d'aquestes esferes.No se sap si aquest procés aparegué abans o després de l'origen del replicador, o potser fou el replicador. La teoria més acceptada és que el replicador, que en aquell moment possiblement era l'àcid ribonucleic (hipòtesi del món d'ARN), juntament amb el seu aparell replicador i possiblement altres molècules, ja havia aparegut.
Les protocèl·lules inicials podrien haver esclatat quan creixien massa; els seus continguts podrien haver recolonitzat altres "bombolles". Les proteïnes que estabilitzaven la membrana o que tenien un paper en la divisió ordenada de les cèl·lules haurien impulsat la proliferació d'aquests llinatges de cèl·lules. L'ARN és un bon candidat pel replicador inicial perquè pot emmagatzemar informació genètica i catalitzar reaccions. En algun moment, l'ADN prengué aquest paper, i les proteïnes conegudes com a enzims passaren a ocupar-se de la catàlisi, deixant que l'ARN transferís informació i controlés el procés. Hi ha la creença, cada cop més estesa, que aquestes cèl·lules primitives podrien haver evolucionat prop de guèisers submarins coneguts com a black smokers.
Nogensmenys, es creu que d'aquesta varietat de cèl·lules, o protocèl·lules, només en sobrevisqué un tipus. Els indicis suggereixen que l'últim avantpassat comú universal visqué a principis de l'eó Arqueà, fa uns 3.500 milions d'anys o encara abans. Aquesta cèl·lula UACU és l'avantpassada de totes les cèl·lules i per tant de tota la vida a la Terra. Probablement es tractava d'un procariota, amb membrana cel·lular i probablement amb ribosomes, però sense nucli ni orgànuls dins la membrana, com serien els mitocondris o els cloroplasts. Com totes les cèl·lules modernes, feia servir l'ADN com a codi genètic, l'ARN per la transmissió d'informació i la síntesi de proteïnes, i enzims per catalitzar les reaccions. Alguns científics creuen que l'últim avantpassat comú universal no era un únic organisme sinó una població d'organismes que s'intercanviaven gens en una transferència horitzontal de gens.
Fotosíntesi i oxigen

És probable que les cèl·lules inicials fossin totes heteròtrofes, utilitzant les molècules orgàniques properes (fins i tot les d'altres cèl·lules) com a matèria primera i font d'energia.[2 A mesura que les fonts alimentàries s'exhauriren, algunes cèl·lules desenvoluparen una nova estratègia. En lloc d'explotar les fonts minvants de molècules orgàniques en estat lliure, aquestes cèl·lules adoptaren la llum solar com a font d'energia. Hi ha diverses estimacions, però fa 3.000 milions d'anys.probablement ja havia aparegut quelcom de semblant a la fotosíntesi moderna. Això féu que l'energia solar pogués ser aprofitada no només pels autòtrofs, sinó també pels heteròtrofs que els consumien. La fotosíntesi utilitzava els abundants diòxid de carboni i aigua com a matèries primeres i, amb l'energia de la llum solar, produïa molècules orgàniques riques en energia (carbohidrats).
A més, la fotosíntesi deixava anar oxigen com a residu. Al principi s'unia amb pedra calcària, ferro i altres minerals. Hi ha proves substancials d'aquest fet en les capes riques en òxids de ferro que corresponen a aquest període. Els oceans s'haurien tornat de color verd a mesura que l'oxigen reaccionava amb els minerals. Quan les reaccions arribaren a la seva fi, l'oxigen començà finalment a acumular-se a l'atmosfera. Malgrat que cada cèl·lula produïa una quantitat ínfima d'oxigen, la combinació del metabolisme de moltes cèl·lules durant un vast període transformà l'atmosfera terrestre en el que és actualment.
Aquesta és, doncs, la tercera atmosfera de la Terra. Una part de l'oxigen fou estimulat per la radiació ultraviolada del Sol i es convertí en ozó, que s'acumulà en una capa a la part alta de l'atmosfera. La capa d'ozó absorbia, i encara absorbeix, una part significativa dels raigs ultraviolats que abans travessaven l'atmosfera. Això permeté a les cèl·lules colonitzar la superfície de l'oceà i, més endavant, la terra ferma: sense la capa d'ozó, el bombardeig continu de raigs ultraviolats hauria causat un nivell insostenible de mutacions en les cèl·lules que hi estiguessin exposades. A part de subministrar grans quantitats d'energia a les formes de vida i aturar la radiació ultraviolada, els efectes de la fotosíntesi tingueren una tercera influència que canvià el món. L'oxigen era tòxic; és probable que gran part de la vida a la Terra s'extingís a mesura que el nivell d'aquest gas creixia (la catàstrofe de l'oxigen). Els organismes resistents sobrevisqueren i prosperaren, i alguns d'ells desenvoluparen la capacitat d'usar l'oxigen per millorar el seu metabolisme i extreure més energia dels mateixos aliments.
6.3. Els pluricel·lulars i la colonització de la Terra
Els arqueobacteris, els eubacteris i els eucariotes
continuaren diversificant-se i tornant-se més sofisticats i més ben
adaptats al seu medi ambient. Cada domini es dividí repetidament en
múltiples llinatges, tot i que no se sap gaire sobre la història dels
arqueobacteris i bacteris. Fa uns 1.100 milions d'anys, el supercontinent de Rodínia s'estava formant. Els llinatges de plantes, animals i fongs
ja s'havien separat, tot i que encara existien com a cèl·lules
solitàries. Algunes d'elles vivien en colònies, i gradualment començà a
aparèixer la divisió de funcions; per exemple, les cèl·lules de la perifèria podrien haver començat a assumir papers diferents dels de les de l'interior.
Fa uns 1.100 milions d'anys aparegueren les primeres plantes pluricel·lulars, probablement cloròfits,
tot i que la línia que separa una colònia amb cèl·lules especialitzades
dels organismes pluricel·lulars no és sempre clara. És probable que fa
uns 900 milions d'anys, la veritable pluricel·lularitat s'hagués desenvolupat també en els animals. Se suposa que al principi era semblant a la de les esponges actuals, en què totes les cèl·lules són totipotents i un organisme danyat es pot regenerar.
A mesura que la divisió de funcions es feia més completa en tots els
llinatges d'organismes pluricel·lulars, les cèl·lules esdevingueren més
especialitzades i més dependents l'una de l'altra; les cèl·lules
aïllades morien.
Molts científics creuen que fa uns 770 milions d'anys començà una època glacial tan severa que es glaçà la superfície de tots els oceans (Terra bola de neu). Finalment, després de 20 milions d'anys, sorgí prou diòxid de carboni de les erupcions volcàniques per a causar un efecte hivernacle que provocà una elevació de les temperatures globals.Al mateix període, fa uns 750 milions d'anys Rodínia començà a fragmentar-se.
L'acumulació d'oxigen a l'atmosfera terrestre portà a la formació d'ozó, que formà una capa que bloqueja la majoria dels rajos ultraviolats que arriben del Sol.
Per això, els organismes unicel·lulars que arribaren a la terra tenien
més possibilitats de sobreviure, i els procariotes començaren a
multiplicar-se i a adaptar-se per a la supervivència fora de l'aigua. És
probable que els procariotes ja haguessin colonitzat la terra fa 2.600
milions d'anys,
abans fins i tot de l'origen dels eucariotes. Durant molt de temps, la
terra restà deserta i sense organismes pluricel·lulars. El
supercontinent Pannòtia es formà fa 600 milions d'anys i es fragmentà només 50 milions d'anys més tard. Els peixos, els vertebrats més antics, evolucionaren als oceans fa uns 530 milions d'anys. Una gran extinció en massa tingué lloc cap a la fi del període Cambrià,que acabà fa 488 milions d'anys.
Fa uns quants centenars de milions d'anys, començaren a créixer plantes (probablement similars a les algues) i fongs a les aigües somes, i més tard fora de l'aigua.Els fòssils més antics de plantes i fongs terrestres daten de fa
480-460 milions d'anys, tot i que anàlisis moleculars suggereixen que
els fongs podrien haver arribat a la terra fa uns 1.000 milions d'anys i
les plantes fa uns 700 milions d'anys.
Al principi romangueren propers a la riba, però una sèrie de mutacions i
variacions els permeté avançar en la colonització d'aquest nou medi. No
es coneix el moment en què els animals abandonaren l'oceà per primer
cop: la prova clara més antiga és l'existència d'artròpodes a la terra fa uns 450 milions d'anys,
que possiblement prosperaven i s'adaptaven millor gràcies a l'enorme
quantitat d'aliments que els proporcionaven les plantes terrestres.
També hi ha indicis no confirmats que els artròpodes podrien haver
arribat a la terra fa 530 milions d'anys.
A finals del període Ordovicià, fa 440 milions d'anys, tingueren lloc més extincions en massa, potser a causa d'una nova època glacial.Més endavant, ara fa uns 380-375 milions d'anys, els primers tetràpodes evolucionaren dels peixos.Es creu que possiblement les aletes es transformaren per convertir-se
en membres que permeteren als primers tetràpodes recolzar-se per treure
els seus caps fora de l'aigua i respirar aire.
Això els permetria sobreviure en aigües pobres en oxigen o perseguir
preses petites a les aigües somes. El següent pas evolutiu permeté que
poguessin sortir a terra ferma durant períodes curts. Finalment, alguns
d'ells s'adaptaren tan bé a la vida terrestre que començaren a passar la
seva vida adulta a la terra, tot i que havien de néixer en el medi
aquàtic i en aquest medi hi seguien realitzant la posta dels ous. Així
aparegueren els amfibis. Fa uns 365 milions d'anys tingué lloc un altre període d'extincions, causat possiblement per un refredament global.Les plantes desenvoluparen llavors, cosa que accelerà dràsticament la seva expansió a la terra durant aquest període.
Uns vint milions d'anys més tard, fa 340 milions d'anys,l'aparició de l'ou amniòtic permeté la posta d'ous a la terra ferma, cosa que suposà un gran avantatge pels embrions dels tetràpodes. Això donà com a resultat la separació entre amniotes i amfibis. Uns trenta milions d'anys després,els sinàpsids (incloent-hi els mamífers) se separaren dels sauròpsids (incloent-hi els ocells i els rèptils
no aviaris i no mamífers). Per descomptat, continuaren evolucionant
altres grups d'organismes i apareixent nous llinatges, però els detalls
no són gaire coneguts. Fa 300 milions d'anys es formà el supercontinent
més recent, Pangea.
L'extinció en massa més gran de la història de la Terra, l'extinció permiana, tingué lloc fa 250 milions d'anys, al límit dels períodes Permià i Triàsic; el 95% de la vida a la Terra s'extingí,possiblement a causa dels trapps siberians, un esdeveniment de gran activitat volcànica.
El descobriment d'un cràter amagat sota la Capa de Glaç de l'Antàrtida
Oriental ha suggerit una nova teoria que un meteorit causà l'extinció i
inicià la fragmentació de Gondwana, creant la falla tectònica que impulsà Austràlia cap al nord.Tanmateix, la vida resistí, i fa uns 230 milions d'anys, els dinosaures
es diferenciaren dels seus avantpassats rèptils. Una extinció a finals
del Triàsic no afectà gaire els dinosaures i fa uns 200 milions d'anys
ja es trobaven en una posició dominant entre els vertebrats. Malgrat
que alguns llinatges de mamífers es començaren a separar durant aquest
període, els mamífers d'aquella època eren amb tota probabilitat petits
animals similars a les musaranyes.
Fa 180 milions d'anys, Pangea es fragmentà en Lauràsia i Gondwana. La frontera entre dinosaures aviaris i no aviaris no és clara, però l'Archaeopteryx, considerat tradicionalment un dels primers ocells, aparegué fa uns 150 milions d'anys. Els indicis més antics de plantes amb flor corresponen al període Cretaci, uns vint milions d'anys després, és a dir, fa uns 132 milions d'anys. La competència de les aus conduí molts gèneres de pterosaure a l'extinció, i els dinosaures ja estaven en declivi per diversos motius quan, fa 65 milions d'anys, un meteorit d'uns 10 quilòmetres de diàmetre impactà amb la Terra just davant de la península del Yucatán, expulsant enormes quantitats de pols i vapor a l'aire, que blocaren la llum solar, evitant la fotosíntesi. La gran majoria d'animals de grans dimensions, incloent-hi els dinosaures no aviaris, s'extingiren, marcant la fi del Cretaci i de l'era Mesozoica.
A partir d'aleshores, durant el Paleocè,
els mamífers no trigaren a diversificar-se, créixer i esdevenir els
vertebrats dominants. L'últim avantpassat comú de tots els primats degué viure uns dos milions d'anys més tard, fa uns 63 milions d'anys.Cap a finals de l'Eocè, fa 40 milions d'anys, alguns mamífers terrestres ja havien tornat als oceans i evolucionat en els avantpassats dels dofins i de les balenes.
Els organismes pluricel·lulars

La colonització de la terra


6.4. Endosimbiosi i els tres dominis de la vida
La taxonomia moderna classifica la vida en tres dominis. El temps de l'origen d'aquests dominis és objecte d'especulació. Probablement, els eubacteris foren els primers a separar-se de les altres formes de vida però aquesta suposició és controvertida. Poc després, fa uns 2.000 milions d'anys, es dividiren en Archaea i Eukarya. Les cèl·lules eucariotes són més grans i complexes que les procariotes (Bacteria i Archaea), i el coneixement que es té de l'origen d'aquesta complexitat és recent. En aquest temps, una cèl·lula bacterial, propera a la Rickettsia actual,s'introduí en una cèl·lula procariota més gran. És possible que la cèl·lula gran intentés digerir la petita però no ho aconseguí, potser a causa de l'aparició de les defenses contra els predadors, o potser la cèl·lula petita intentà parasitar la gran; fos com fos, sobrevisqué a l'interior de la gran.
La cèl·lula petita, utilitzant oxigen, fou capaç de metabolitzar els residus de la cèl·lula gran i extreure'n més energia, de la qual una part tornava a l'hostatjador. La cèl·lula petita es replicà dins la gran, i aviat es desenvolupà una relació simbiòtica estable. Amb el temps, l'hostatjador adquirí alguns gens de les cèl·lules més petites, i els dos tipus d'organisme es tornaren interdependents: la cèl·lula gran no podia sobreviure sense l'energia produïda per les petites, i aquestes no podien sobreviure sense la matèria primera que els subministrava la cèl·lula gran. Aquesta simbiosi entre la cèl·lula gran i les cèl·lules petites del seu interior es desenvolupà fins al punt que es considera que es convertiren en un únic organisme, amb les cèl·lules petites classificades com a orgànuls amb el nom de mitocondris.
Passà quelcom de semblant amb els cianobacteris fotosintètics que s'introduïren dins cèl·lules heteròtrofes més grans, convertint-se en cloroplasts. Probablement com a resultat d'aquests canvis, un llinatge de cèl·lules
amb capacitat de fotosíntesi se separà dels altres eucariotes fa un xic
més de 1.000 milions d'anys. Segurament hi hagué diverses instàncies
d'aquest fenomen, com es veu a la imatge de la dreta. A part de la
teoria endosimbiòtica de l'origen cel·lular dels mitocondris i
cloroplasts, àmpliament acceptada, s'ha suggerit que les cèl·lules
portaren als peroxisomes, les espiroquetes als cilis i flagels, i que un virus ADN podria haver donat peu al nucli cel·lular tot i que cap d'aquestes teories no s'ha guanyat molta acceptació.
6.5. Éssers humans
Un petit primat africà que visqué fa uns sis milions d'anys fou l'últim avantpassat comú dels humans actuals i els seus parents més propers, els bonobos i ximpanzés. Només dues de les branques d'aquest arbre evolutiu no s'han extingit. Poc després de la separació, i per raons que encara són debatudes, els primats d'una de les branques desenvoluparen l'habilitat de caminar dempeus.La mida del cervell augmentà ràpidament, i fa uns 2 milions d'anys ja havien aparegut els primers animals classificats en el gènere Homo.Evidentment, la línia que separa diferents espècies o fins i tot gèneres és bastant arbitrària, car els organismes canvien contínuament amb cada generació.
Durant el mateix període, l'altra branca es dividí en els avantpassats del ximpanzé comú i els avantpassats del bonobo, mentre que l'evolució continuava simultàniament en totes les formes de vida.És probable que els primers a manipular el foc fossin els Homo erectus (o Homo ergaster), fa almenys 790.000 anys, però podria ser que es remuntés a fa 1,5 milions d'anys. És més difícil establir l'origen del llenguatge; no està clar si Homo erectus podria parlar o si Homo sapiens fou el primer a aconseguir-ho.
A mesura que la mida del cervell creixia, les cries naixien més aviat, abans que els seus caps fossin massa grans per passar per la pelvis. Per tant, mostraven més neuroplasticitat i posseïen una capacitat especial d'aprendre, però eren dependents dels pares durant més temps. Les habilitats socials es tornaren més complexes, el llenguatge més avançat i les eines més elaborades. Això portà a encara més cooperació i a un major desenvolupament cerebral.
Es creu que els humans anatòmicament moderns, Homo sapiens, s'originaren fa uns 200.000 anys a Àfrica; els fòssils més antics daten de fa aproximadament 160.000 anys.Els primers humans que presenten proves d'espiritualitat són els neandertals, sovint classificats com a una espècie diferent que s'extingí; enterraven els morts, generalment amb menjar o eines. Nogensmenys, no aparegueren proves de cultes més sofisticats fins fa uns 32.000 anys,amb les pintures rupestres dels cromanyons, que probablement tenien un significat màgic o religiós.Els cromanyons també deixaren figures de pedra com ara la Venus de Willendorf, probablement una estàtua amb un significat religiós. Fa 11.000 anys, Homo sapiens ja havia arribat a l'extrem meridional d'Amèrica del Sud, l'últim dels continents no habitats, amb l'excepció de l'Antàrtida.L'ús d'eines i el llenguatge continuaren perfeccionant-se, i les relacions interpersonals esdevingueren més complexes.
Quadre resum:
5 milions d'anys
|
Els avantpassats dels humans s'especien dels avantpassats mutus dels ximpanzés. L'últim avantpassat comú és el Sahelanthropus tchadensis. El més primerenc en la branca humana és l'Orrorin tugenensis. Tant els ximpanzés com els humans tenen una laringe que es ressitua durant els dos primers dos anys de vida en un punt entre la faringe i els pulmons, indicant que els ancestres comuns tenien aquesta característica precursora de la parla. |
| 4,4 milions d'anys | Ardipithecus ramidus ramidus (Potser caminava dempeus la major part del temps o potser encara era més arborícola que terrícola) |
| 3,7 milions d'anys | Alguns Australopithecus afarensis deixen les seves petjades en cendres volcàniques de Laetoli a Kenya. |
| 3 milions d'anys | Els bípedes autralopitecins (hominins primerencs) es desenvolupen a les sabanes d'Àfrica. |
2 milions d'anys
|
Homo habilis es creu que és l'avantpassat de Homo ergaster, que al mateix temps va doanr lloc a l'espècie més humana de Homo erectus. Hi ha un debat sobre si H. habilis és un avantpassat directe i sobre quants dels fòssils coneguts han de ser atribuïts a aquesta espècie.
|
| 1,8 milions d'anys |
Homo erectus apareix a l'Àfrica. Tindria una semblança evident amb els homes moderns però el seu cervell només seria un 74 per cent de la mida dels humans moderns. El seu front està menys inclinat i les dents són més petites. Es creu que és un avantpassat dels humans moderns (considerant Homo heidelbergensis com a un pas intermediari). |
| 1,75 milions d'anys | Homo georgicus (Geòrgia), de petit cervell, s'hi desplaça des d'Àfrica amb característiques de Homo erectus i Homo habilis. |
| 700 milers d'anys | Avantpassat genètic comú dels humans moderns i els neanderthals. |
| 355 milers d'anys | Un Homo heidelbergensis de 1,5 m d'alçada deixa petjades en cendres volcàniques solidificades d'Itàlia. És morfològicament molt semblant a Homo erectus però té un major cervell que fa un 93% de la mida de Homo sapiens. L'espècie era alta de mitjana (1,8 m) i més musculada que els humans moderns. |
| 195 milers d'anys | Omo1, Omo2 (Etiòpia, riu Omo) són els primers Homo sapiens coneguts. |
| 160 milers d'anys | L'Homo sapiens (Homo sapiens idaltu) a Etiòpia, al riu Awash, a Herto, practica rituals funeraris. |
| 150 milers d'anys | Naixement de l'Eva mitocondrial a l'Àfrica. És la dona més recent avantpassada de la totalitat d'humans actualment vius. |
| 130 milers d'anys | Apareix el gen relacionat amb el desenvolupament de la parla. |
100 milers d'anys
|
Els primers éssers humans anatòmicament moderns (Homo sapiens) apareixen a Àfrica, evolucionant de Homo heidelbergensis. Segons les estimacions actuals, els éssers humans tenen aproximadament 20.000–25.000 gens i compartien el 99% del seu ADN amb espècies extintes de Neanderthals i un 95% amb els ximpanzés, els parents evolutius vius més propers. La pell de Homo sapiens està relativament mancada de pèl comparada amb la dels altres primats. El color de pell dels éssers humans contemporanis pot anar des del marró molt fosc fins al rosa pàl·lid. Està geogràficament estratificat i generalment es correspon amb els nivells d'UV. La pell humana i el color de cabell està controlada en part pel gen MC1R. Per exemple, el pèl roig i la pell pàl·lida d'alguns europeus és el resultat de la mutació en el gen MC1R. La pell humana té la capacitat de broncejar-se en resposta a l'exposició dels UV. La variació en la capacitat de bronzejar-se està també controlada pel gen MC1R. |
| 90 milers d'anys | Els humans moderns arriben a l'Àsia a través de dues rutes: una pel nord, a través del Pròxim Orient i una pel sud a través d'Etiòpia, a través del Mar Roig i el sud d'Aràbia. Les mutacions causen canvis de color per tal d'absorbir les quantitats òptimes d'UV per a diferents latituds geogràfiques. Es comencen a formar les diferents "races" modernes. Les poblacions africanes romanen genèticament més diverses que la resta d'humanitat junta perquè només un subgrup de la seva població (i per tant només un subgrup de la seva població) va abandonar l'Àfrica. |
| 74 milers d'anys | Erupció supervolcànica de Toba, Sumatra, Indonèsia fa disminuir la població humana. 6 anys d'hivern nuclear i 1000 anys d'Edat de Gel. |
| 60 milers d'anys | Naixement del cromosoma Y Adam a l'Àfrica (probablement a Etiòpia o al Sudan). És l'ancestre comú més recent del que descendeixen tots els cromosomes Y humans. |
| 50 milers d'anys | Els humans moderns s'expandeixen des d'Àsia fins a Austràlia (per esdevenir els aborígens) i també a Europa. L'expansió es realitza més ràpidament per les costes que pels interiors. |
| 31 milers d'anys | Els humans moderns entren per Sibèria a Amèrica del Nord
en diverses onades. Com a mínim dues de les primeres onades no han
deixat rastre genètic a Amèrica actualment. Els humans també arriben a
les illes Salomó i al Japó.
Apareix el marcador genètic M343. Es creu que va aparéixer en un home d'Àfrica fa uns 30.000 o més anys, propagant-se des de llavors. Aquest marcador genètic el porten la majoria d'europeus occidentals. El porten el 70% de la població total d'Anglaterra i el 90% en algunes part d'Irlanda i de la península Ibèrica. |
| 27 milers d'anys | Els neanderthals s'extingeixen deixant Homo sapiens com l'única espècie viva del gènere Homo. |
| 10 milers d'anys | Els humans arriben a la Terra del Foc a l'extrem sud de l'Amèrica del Sud, l'última regió continental en ser habitada per humans (excloent-ne l'Antàrtida). |
Per saber-ne més: https://www.wikiwand.com/ca/Evoluci%C3%B3_dels_%C3%A9ssers_humans
7. Resum sobre lamarquisme, darwinisme i neodarwinisme
La teoria de Lamarck : el lamarquisme
Jean Baptiste de Lamarck (1769-1832) fou un famós naturalista. Va formular la primera teoria coherent sobre el mecanisme a través del que es transformen els organismes. Es va basar en 2 principis :
- la capacitat dels organismes per adaptar-se a les circumstàncies (a l’ambient),
- l’herència dels caràcters adquirits.
Avui ja s’ha demostrat la falsedat de tots 2 principis, però alguns d’ells constituïen fermes creences en l’època de Lamarck.
Aquests són els fonaments de la teoria de Lamarck :
1 A mesura que les circumstàncies de territori, exposició al sol, clima, alimentació, hàbits de vida, etc., canvien,els caràcters de mida, forma, d’agilitat i d’activitat, dels animals i plantes, canvien proporcionalment.
2 La falta d’ús d’un òrgan, feta constant pels hàbits adquirits, empobreix gradualment aquest òrgan i acaba per fer-lo desaparèixer. (Per exemple, tenir ulls al cap és propi de gran nombre d’animals diversos; formen una part essencial del pla d’organització dels vertebrats. No obstant el talp, que pels seus hàbits fa servir poc la vista, té uns ulls molt petits gairebé no són aparents, perquè exercita molt poc aquests òrgans).
3 L’ús freqüent d’un òrgan, fet constant pels hàbits, augmenta la facultat d’aquest òrgan, el fa desenvolupar i el fa adquirir unes mides i una força d’acció que no es poden comparar ni de lluny amb els animals que l’exerciten menys. (Un altre exemple: Aquella au que per necessitat ha d’acudir a l’aigua per trobar la presa que el permeti viure, separa els dits de les potes quan vol batre l’aigua i moure’s per la superfície. La pell que uneix aquests dits per la base desenvolupa com a conseqüència l’hàbit d’estendre’s. Així, amb el temps, es formen tal com les veiem, les grans membranes que uneixen els dits dels ànecs, oques, etc).
4 Tot canvi adquirit en un òrgan per un hàbit d’ús suficient per haver-lo produït es conserva per la generació si el canvi és comú als individus que, en la fecundació, intervenen en la reproducció de l’espècie. El canvi es propaga i passa així a tots els individus que es succeeixen i que estan sotmesos a les mateixes circumstàncies, sense que calgui adquirir-lo per la via que realment el va crear.
PRIMERA LLEI
En tot animal que no hagi sobrepassat el final del seu desenvolupament,l’ús més freqüent i sostingut d’un òrgan qualsevol fortifica poc a poc aquest òrgan,el desenvolupa, l’engrandeix i li dóna una potència proporcional a la durada de la seva utilització. En canvi la falta constant d’ús d’un òrgan, el debilita de forma inapreciable,el deteriora, disminueix progressivament les seves facultats i acaba per fer-lo desaparèixer.
SEGONA LLEI
Tot el que la naturalesa ha fet adquirir o perdre als individus per efecte de les circumstàncies a que ha estat exposada la seva espècie durant llarg temps i, en conseqüència, a causa de l’ús predominant de l’esmentat òrgan o d’una falta constant de la seva utilització, és conservat en els individus descendents.
La Teoria de la selecció natural: el darwinisme
La teoria de la selecció natural va ser proposada per Charles Darwin (1809-1882). Possiblement el viatge que Darwin va fer de jove al voltant del món va despertar en ell l’interès per la diversificació dels organismes i pel procés que origina les espècies. L’observació de la flora i fauna de Sudamèrica, i especialment de les Illes Galápagos, fou decisiva en l’elaboració de la teoria de la selecció natural. Aquesta teoria es va publicar el 24 de novembre de 1859 en el llibre anomenat L’origen de les espècies. Havien passat més de 20 anys, dedicats a l’estudi i al aprofundiment en les seves idees, des del seu viatge al voltant del món.
Analitzem ara els principals enunciats de la Teoria de la selecció natural.
1 Les poblacions d’organismes tenen un potencial reproductor que les pot conduir a un creixement exponencial. Però, en canvi,en períodes no massa llargs de temps el número d’individus d’una població es manté aproximadament constant.
2 Això es degut a que les condicions de vida limiten el creixement de les poblacions. L’entorn (característiques ambientals) dels organismes ha experimentat canvis constants a través del temps geològic.
3 Només una part de la descendència d’una població viu per a produir descendència (no totes les cries arriben a adults, no tots els adults arriben a reproduir-se). Aquest fenomen és la lluita per a l’existència, els organismes han de superar les condicions que els imposa el medi on viuen.
4 Els individus d’una població no són tots iguals,mostren variacions que són heretables (característiques diferencials).
5 El medi imposa unes condicions difícils per a la vida dels organismes. Segons com sigui aquest medi, algunes característiques dels organismes seran favorables (permetran desenvolupar activitats que proporcionen èxit,arribar a adult i deixar descendència), mentre que d’altres seran desfavorables (ser portador d’aquella característica representa un perjudici per arribar a adult o produir descendència).
6 Els individus portadors de característiques favorables produiran més descendència que altres individus que no tenen aquestes característiques; en conseqüència els individus que són portadors de característiques desfavorables produeixen menys descendència en relació a la resta de la població.
La població, anirà canviant poc a poc, generació a generació. Es diu que pateix un procés d’adaptació, d’acomodació gradual al medi on viu.
7 La selecció natural causarà, després de moltes generacions, l’acumulació de característiques favorables (en relació al medi en què els organismes viuen) i la desaparició de les característiques desfavorables.
El Neodarwinisme admet postulats del Darwinisme però afegint coneixements de la genètica. Per això refusa radicalment l’herència dels caràcters adquirits i considera la població (i no l’individu) com la unitat evolutiva.
Podem resumir el neodarwinisme en els següents punts:
a) Les mutacions són una font primària de variabilitat genètica i la recombinació genètica augmenta aquesta variabilitat.
b) La variabilitat genètica es tradueix en nous fenotips.
c) Les variants al·lèliques tenen diferent eficàcia biològica o èxit reproductiu.
d) La selecció natural actua sobre la variabilitat genètica, de manera, que generació rere generació, es modifiquen les freqüències al·lèliques.
e) Els caràcters adquirits no s'hereten.
f) La unitat d'evolució és la població.
8. Exemple de selecció natural i tolerància a la lactosa
Vegeu aquí com influeix l'ambient en la selecció d'unes determinades característiques sorgides per una mutació a l'atzar en una població:
https://www.youtube.com/watch?v=nE_1TcdjdV8&feature=youtu.be
9. Un altre exemple de selecció natural : coevolució
La coevolució és el canvi evolutiu entre dues o més espècies que interactuen, de manera recíproca i gràcies a la selecció natural.
Perquè hi hagi coevolució s’ha de complir:
- Especificitat: l’evolució de cada caràcter d’una espècie es deu a pressions selectives del caràcter de l’altra espècie.
- Reciprocitat: els caràcters evolucionen de manera conjunta.
- Simultaneïtat: els caràcters evolucionen al mateix temps.
Ja sabem que és inevitable que els éssers vius estableixen relacions de simbiosi entre ells. Uns depenen d’altres per sobreviure, i alhora, de l’accés a elements del seu entorn com aigua, llum o aire. Aquestes pressions mútues entre espècies fan que evolucionin conjuntament i segons evolucioni una espècie, obligarà al seu torn a l’altra a evolucionar. Vegem alguns exemples:
POL·LINITZACIÓ
El procés més conegut de coevolució el trobem en la pol·linització.
Els insectes ja existien molt abans de l’aparició de plantes amb flor, però el seu èxit es va deure al descobriment que el pol·len és una bona reserva d’energia. Al seu torn, les plantes troben en els insectes una manera més eficaç de transportar el pol·len cap a una altra flor. La pol·linització gràcies al vent requereix més producció de pol·len i una bona dosi d’atzar perquè almenys algunes flors de la mateixa espècie siguin fecundades. Moltes plantes han desenvolupat flors que atrapen als insectes fins que estan coberts de pol·len i els deixen escapar. Aquests insectes presenten pèls en el seu cos per permetre aquest procés. Al seu torn alguns animals han desenvolupat llargs apèndixs (becs dels colibrís, espiritrompes de certes papallones…) per accedir al nèctar.

10. Errades freqüents
Errades freqüents
El tema de l'evolució és senzill però a la vegada ple de malentesos ja que fem servir conceptes que no entenem bé o els fem servir malament. Aquí us poso errades detectades.
1. A les poblacions hi ha individus diferents. Tenen genotips diferents. Què origina la variabilitat de les espècies?
Com que molts caràcters que tenim són heretables, la variabilitat genètica és deguda a les mutacions (canvis en l'ADN) i a la recombinació que es produeix en la reproducció sexual (recordeu que quan es formen els gàmetes a la meiosi hi ha recombinació). Important: Aquests processos són a l'atzar (aleatoris). No hi ha cap força que empenyi a canviar en una determinada direcció... (Alguns pensen que els individus s'esforcen per adaptar-se a les característiques del medi i per això muten. FALS). Les mutacions són preadaptatives. Això vol dir que són les que són i poden ser favorables, desfavorables o neutres.
2. No hem de confondre selecció i mutació. Què és la selecció natural?
La mutació hem dit que és font de variabilitat. La selecció és el procés que fa que si una mutació és idònia en un medi determinat, sigui seleccionada (afavorida) perquè en aquell medi , els que la tinguin, sobreviuran més i es podran reproduir més. La selecció fa que els genotips més favorables deixin més descendència i per això la freqüència de determinats gens augmenta a la població.
3. No hem de creure en idees lamarckianes. Avui dia estan descartades.Cap evidència científica no els dona suport.
Els canvis deguts a l'ús i desús dels òrgans NO s'hereten ( un jugador de futbol amb cames musculades no tindrà un fill que les hi tingui si aquest no fa aquest esport, per exemple)
El medi no origina necessitats noves. (per exemple, uns animals que viuen a l'aigua, no necessiten adaptar-s'hi; això és incorrecte dit així. El correcte és dir que en el medi aquàtic, els que tinguin característiques favorables- originades per mutacions- sobreviuran millor i es reproduiran més, transmetent aquests caràcters als descendents).
Agafem, per exemple, el cas del gran desenvolupament del coll de les girafes. Segons el lamarckisme, les girafes tenen el coll molt llarg perquè generació rere generació han anat estirant el coll per tal d'arribar a les fulles dels arbres dels quals s'alimenten. El fet d'estirar el coll provoca que s'allargui una mica. Aquest petit canvi es pot heretar, de manera que la descendència té ja de bon començament el coll una mica més llarg. Si això es va repetint durant moltes generacions, s'aconsegueix que el coll esdevingui molt llarg. AIXÒ NO ÉS CORRECTE.
Segons el darwinisme, originàriament hi havia girafes amb el coll més llarg i girafes amb el coll més curt . En èpoques de sequera o de molta competència pel menjar, les girafes amb el coll més llarg podien arribar a les fulles més altes i per tant tenien accés a més menjar. Les girafes amb el coll més curt no podien arribar a les fulles més altes i morien de fam . Això feia que només es poguessin reproduir les girafes de coll llarg i, per tant, a la següent generació les girafes tendissin a tenir el coll més llarg . Tanmateix, hi continuaria havent variabilitat de mides de coll, per la qual cosa el procés de selecció es podria continuar produint. Després de moltes generacions, la selecció natural aconseguiria que el coll de les girafes fos molt més llarg que no pas originàriament.